Los lípidos son compuestos orgánicos insolubles en agua (o sólo poco solubles), que se
encuentran en los sistemas biológicos. Los lípidos tienen gran solubilidad en solventesorgánicos no polares. Son hidrofóbicos (no polares) o bien son anfipáticos (contienen regiones polares y no polares al mismo tiempo). Los lípidos más simples son los ácidos grasos, y tienen la fórmula general R—COOH, donde R representa una cadena de hidrocarburo.
Los ácidos grasos son componentes de muchos tipos más complejos de lípidos, incluyendo los triglicéridos o triacilgliceroles, los glicerofosfolípidos y los esfingolípidos. Los lípidos que contienen grupos fosfato se llaman fosfolípidos y los que tienen grupos esfingosina y carbohidrato a la vez se llaman glicoesfingolípidos. Los esteroides, las vitaminas lipídicas y los terpenos se relacionan con la molécula de isopreno, de cinco carbonos, y por consiguiente se llaman isoprenoides. El nombre terpenos se ha aplicado a todos los isoprenoides, pero en general se restringe a los que existen en las plantas.

Ácidos grasos
 |
| Nomenclatura de los ácidos Grasos |
Los ácidos grasos difieren entre sí en la longitud de sus colas de hidrocarburo, la cantidad de dobles enlaces carbono-carbono, las posiciones de los dobles enlaces en las cadenas y la cantidad de ramificaciones. Son una forma de detergente, porque tienen una larga cola hidrofóbica y una cabeza polar. La cantidad de átomos de carbono en los ácidos grasos más abundantes va de 12 a 20, y casi siempre es par, ya que se sintetizan mediante la adición consecutiva de unidades con dos carbonos. En la nomenclatura común se usan letras griegas para identificar a los átomos de carbono. El carbono adyacente al carbono carboxílico (C-2, en la nomenclatura de IUPAC) se le denomina a, y los demás carbonos tienen las letras b, g, d, e, etcétera.La letra griega w (omega) especifica el átomo de carbono más alejado del grupo carboxilo, cualquiera que sea la longitud de la cola de hidrocarburo. (w es la última letra del alfabeto griego).
 Los ácidos grasos que no contienen dobles enlaces carbono-carbono se llaman saturados, en tanto que los que tienen al menos un doble enlace carbono-carbono se clasifican como no saturados o insaturados. Los ácidos grasos no saturados que sólo tienen un doble enlace carbono-carbono se llaman monoinsaturados, en tanto que los que tienen dos o más se denominan poliinsaturados. La configuración de los dobles enlaces en los ácidos grasos no saturados es cis, en general.
Los ácidos grasos que no contienen dobles enlaces carbono-carbono se llaman saturados, en tanto que los que tienen al menos un doble enlace carbono-carbono se clasifican como no saturados o insaturados. Los ácidos grasos no saturados que sólo tienen un doble enlace carbono-carbono se llaman monoinsaturados, en tanto que los que tienen dos o más se denominan poliinsaturados. La configuración de los dobles enlaces en los ácidos grasos no saturados es cis, en general.
Ejemplo de ácidos grasos:
Las propiedades físicas de los ácidos grasos saturados y no saturados son muy variadas. Típicamente, los ácidos grasos saturados son sólidos céreos a temperatura ambiente (22 °C), en tanto que los ácidos grasos no saturados son líquidos a esta temperatura. La longitud de la cadena de hidrocarburo en un ácido graso, y su grado de insaturación, influyen sobre el punto de fusión

Estructura de los principales ácidos grasos poliinsaturados

Triacilgliceroles o Triglicéridos
Son combustibles metabólicos importantes, en especial en los mamíferos.
La oxidación de los ácidos grasos produce mas energía (37 kJ g-1) que la oxidación de proteínas y carbohidratos (16 kJ g-1 cada uno).
Los ácidos grasos se almacenan en forma de lípidos neutros llamados triacilgliceroles.
Los triacilgliceroles están formados por tres residuos de acilo graso esterificados con glicerina.
Los triacilgliceroles son muy hidrofóbicos. Se almacenan en células en forma anhidra, las moléculas no están solvatadas por el agua.
Son combustibles metabólicos importantes, en especial en los mamíferos.
La oxidación de los ácidos grasos produce mas energía (37 kJ g-1) que la oxidación de proteínas y carbohidratos (16 kJ g-1 cada uno).
Los ácidos grasos se almacenan en forma de lípidos neutros llamados triacilgliceroles.
Los triacilgliceroles están formados por tres residuos de acilo graso esterificados con glicerina.
Los triacilgliceroles son muy hidrofóbicos. Se almacenan en células en forma anhidra, las moléculas no están solvatadas por el agua.
Las grasas y los aceites son mezclas de triacilgliceroles. Pueden ser sólidos (grasas) o líquidos (aceites), dependiendo de sus composiciones de ácidos grasos y de la temperatura. Los triacilgliceroles que sólo contienen grupos acilo graso saturado y de cadena larga tienden a ser sólidos a la temperatura corporal, y los que contienen grupos acilo graso no saturados o de cadena corta, tienden a ser líquidos.
La mayor parte de los lípidos en la dieta humana promedio son triacilgliceroles. Esos lípidos se descomponen en el intestino delgado por acción de las lipasas.
La palmitina contiene tres residuos de acido palmítico.

La trioleina contiene tres residuos de acido oleico.
Glicerofosfolípidos
Son los lípidos mas abundantes en la mayor parte de las membranas
biológicas.
Los glicerofosfolípidos o fosfogliceridos al igual que los triacilgliceroles tienen
un soportarte de glicerol.
Los fosfatidatos están presentes en pequeñas cantidades como intermedios
en la biosíntesis y descomposición de glicerofosfolípidos.
Son los lípidos mas abundantes en la mayor parte de las membranas
biológicas.
Los glicerofosfolípidos o fosfogliceridos al igual que los triacilgliceroles tienen
un soportarte de glicerol.
Los fosfatidatos están presentes en pequeñas cantidades como intermedios
en la biosíntesis y descomposición de glicerofosfolípidos.

Estructura de algunos glicerofosfolipidos: fosfatidilcolina, fosfatidiletanolamina, fosfatidilserina, fosfatidilinositol y difosfotidilglicerol.

La fosfatidilcolina es una familia de licerofosfolipidos y es el fosfolípido más
abundante en las membranas de las células tanto animales como vegetales donde se encuentra en la cara externa de la membrana.
También forma parte de las lipoproteínas plasmáticas, es fuente de 1,2-diacilglicerolabundante en las membranas de las células tanto animales como vegetales donde se encuentra en la cara externa de la membrana.
(molécula de señalización) mediante la acción de la fosfolipasa D y aporta la fosforilcolina
para la síntesis de esfingomielina.
En general, los glicerofosfolipidos tienen ácidos grasos saturados esterificados en el C-1 y
ácidos grasos no saturados esterificados en el C-2.
La fosfatidiletanolamina es también un componente básico de la cara interna de las membranas, en animales y plantas, puede actuar como carabina molecular durante el ensamblaje de las proteínas de membrana guiando su plegamiento y facilitando su transición desde el entorno del citoplasma a la membrana plasmática.
ácidos grasos no saturados esterificados en el C-2.
La fosfatidiletanolamina es también un componente básico de la cara interna de las membranas, en animales y plantas, puede actuar como carabina molecular durante el ensamblaje de las proteínas de membrana guiando su plegamiento y facilitando su transición desde el entorno del citoplasma a la membrana plasmática.
La fosfatidilserina contribuye a las interacciones electrostáticas no
específicas de la cara interna de las membranas aunque esta localización se puede ver alterada durante la activación de plaquetas o en la apoptosis.
Cuando se transfiere a la cara externa de la membrana y actúa como una señal para otras células.
Junto al diacilglicerol y a los iones Ca2+, es necesaria para la activación de la proteína quinasa C (PKC), una enzima clave en la transducción de señales.
específicas de la cara interna de las membranas aunque esta localización se puede ver alterada durante la activación de plaquetas o en la apoptosis.
Cuando se transfiere a la cara externa de la membrana y actúa como una señal para otras células.
Junto al diacilglicerol y a los iones Ca2+, es necesaria para la activación de la proteína quinasa C (PKC), una enzima clave en la transducción de señales.
Para determinar las estructuras de los glicerofosfolípidos y las identidades de sus ácidos grasos individuales se utilizan diversas fosfolipasas que catalizan en forma específica la hidrolisis de los enlaces éster.
La fosfolipasa A2 es la principal enzima en le jugo pancreático y es la responsable de la
digestión de fosfolípidos de membrana la dieta. También esta presente en los venenos de víboras, abejas y avispas.
digestión de fosfolípidos de membrana la dieta. También esta presente en los venenos de víboras, abejas y avispas.

PlasmalogenosOtra clase principal de glicerofosfolípidos, presenta un sustituyente hidrocarburo en el grupo hidroxilo del C-1 de la glicerina unido por un enlace de éter vinílico y no enlace éster.
La etanolamina o la colina se suelen esterificar al grupo fosfato de los plasmalogenos.Estos forman un 23 % de los glicerofosfolípidos en el sistema nervioso central humano y también se encuentran en las membranas de los tejidos nerviosos periféricos y muscular.
La etanolamina o la colina se suelen esterificar al grupo fosfato de los plasmalogenos.Estos forman un 23 % de los glicerofosfolípidos en el sistema nervioso central humano y también se encuentran en las membranas de los tejidos nerviosos periféricos y muscular.

Esfingolípidos
Después de los glicerofosfolípidos, los lípidos mas abundantes en las membranas vegetales y animales son los esfeingolípidos.
Se encuentra en mamíferos en especial abundancia en el tejido nervioso central. La mayor parte de las bacterias no tienen esfingolipiodos.
Después de los glicerofosfolípidos, los lípidos mas abundantes en las membranas vegetales y animales son los esfeingolípidos.
Se encuentra en mamíferos en especial abundancia en el tejido nervioso central. La mayor parte de las bacterias no tienen esfingolipiodos.
El componente estructural de los esfingolipidos es la esfingosina (trans 4- esfingenina). Es un alcohol no ramificado de C 18, con un doble enlace
trans entre el C-4 y C-5 un grupo amino en el C-2 y grupos hidroxilos en el C-1 y C-3.
trans entre el C-4 y C-5 un grupo amino en el C-2 y grupos hidroxilos en el C-1 y C-3.

Ceramida
La ceramida esta formada por un grupo acilo graso unido al grupo amino del C-2 en la esfingosina, por un enlace amida.
Son los precursores metabólicos de todos los esfingolípidos.
Las tres grandes familias de esfingolípidos son las esfingomielinas, los cerebrosidos y los
gangliosidos.
Solo las esfingomielinas contienen fosfato y se clasifican como fosfolípidos, los cerebrocidos y los gangliosidos contienen residuos de carbohidratos y se clasifican como glicoesfingolípidos.
La ceramida esta formada por un grupo acilo graso unido al grupo amino del C-2 en la esfingosina, por un enlace amida.
Son los precursores metabólicos de todos los esfingolípidos.
Las tres grandes familias de esfingolípidos son las esfingomielinas, los cerebrosidos y los
gangliosidos.
Solo las esfingomielinas contienen fosfato y se clasifican como fosfolípidos, los cerebrocidos y los gangliosidos contienen residuos de carbohidratos y se clasifican como glicoesfingolípidos.

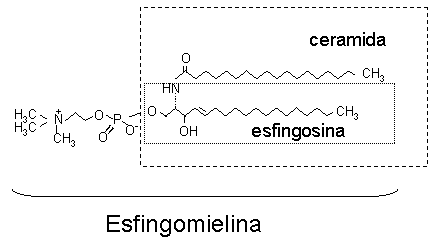
Esfingomielina
La fosfocolina esta unida al grupo hidroxilo en el C-1 de una ceramida
Las esfingomielinas existen en las membranas plasmáticas de la mayor parte de las células de mamíferos y son los componentes principales de las vainas de mielina que rodean a ciertas células nerviosas.
La fosfocolina esta unida al grupo hidroxilo en el C-1 de una ceramida
Las esfingomielinas existen en las membranas plasmáticas de la mayor parte de las células de mamíferos y son los componentes principales de las vainas de mielina que rodean a ciertas células nerviosas.
Cerebrosidos
Galactocerebrosiado
Son esfingolípidos que contienen un residuo de monosacárido unido a un enlace β-glicosídico al C-1 de una ceramida.
Los galactocerebrosidos, llamados también galactosilceramidas, tienen un solo residuo de β-D-galactosilo como grupo de cabeza polar.
Los galactocerebrosidos abundan en el tejido nervioso y forman casi el 15% de los lípidos en la vaina de mielina.
Muchos otros tejidos en los mamíferos contienen glucocerebrosidos, ceramidas con un grupo β-D-glucosilo en la cabeza.
Galactocerebrosiado
Son esfingolípidos que contienen un residuo de monosacárido unido a un enlace β-glicosídico al C-1 de una ceramida.
Los galactocerebrosidos, llamados también galactosilceramidas, tienen un solo residuo de β-D-galactosilo como grupo de cabeza polar.
Los galactocerebrosidos abundan en el tejido nervioso y forman casi el 15% de los lípidos en la vaina de mielina.
Muchos otros tejidos en los mamíferos contienen glucocerebrosidos, ceramidas con un grupo β-D-glucosilo en la cabeza.

Gangliosidos
Son glicoesfingolípidos mas complejos, donde las cadenas de oligosacáridos que contienen ácido N-acetilneuraminico (NeuNAc) están unidos a una ceramida.
Gangliosido GM2, la M representa monosialo, es decir un residuo de NeuNAc y el subíndice 2 ya que es el segundo glangliosido que se caracterizo.
Se han caracterizado mas de 60 variedades de gangliosidos.
Se han caracterizado mas de 60 variedades de gangliosidos.

 Esteroides
EsteroidesEs la tercera clase de lípidos que se encuentran en las membranas de los eucariotas y muy rara vez en bacterias.
Los esteroides junto con las vitaminas lipídicas y los terpenos, se clasifican como isoprenoides, por que su molécula se relaciona con le isopreno.
a) Estructura química
b) Esqueleto de carbono
c) Unidad de isopreno
Los esteroides contiene cuatro anillos fundidos,tres de seis carbonos idénticos como A, B y C y un anillo D de cinco carbonos.
La estructura anular característica se deriva del ESCUALENO
Colesterol
El colesterol es un esterol (lípido) que se encuentra en los tejidos corporales y en el plasma sanguíneo de los vertebrados. Se presenta en altas concentraciones en el hígado, médula espinal, páncreas y cerebro.
Pese a tener consecuencias perjudiciales en altas concentraciones, es esencial para crear la membrana plasmática que regula la entrada y salida de sustancias queatraviesan la célula.
Abundan en las grasas de origen animal.
El colesterol es un esterol (lípido) que se encuentra en los tejidos corporales y en el plasma sanguíneo de los vertebrados. Se presenta en altas concentraciones en el hígado, médula espinal, páncreas y cerebro.

Hay muchas clases de lípidos que no se encuentran en las membrana.Entre ellos tenemos las ceras, eicosanoides y algunos isoprenoides.
Los lípidos no constituyentes de membranas tienen diversas funciones especializadas (ej. Las vitaminas lipídicas).
Los lípidos no constituyentes de membranas tienen diversas funciones especializadas (ej. Las vitaminas lipídicas).

Ceras
Son ésteres no polares de ácidos grasos de cadena larga y alcoholes monohidroxílicos de cadena larga. Las ceras están muy distribuidas en la naturaleza. Proporcionan cubiertas protectoras impermeables a las hojas y frutos de ciertas plantas, y en la piel, cuero, plumas y exoesqueletos de animales. Por ejemplo, el palmitato de miricilo, uno de los principales componentes de la cera de abejas, es el éster de palmitato (16:0) y del alcohol miricílico, de 30 carbonos (figura 9.17). La hidrofobicidad del palmitato de miricilo hace que la cera de abejas sea muy insoluble en agua, y su alto punto de fusión (debido a las cadenas largas de hidrocarburo saturado) hace que esa cera sea dura y sólida a las temperaturas peculiares de la intemperie.
 |
| Palmitato de miricilo |
Los eicosanoides son derivados oxigenados de ácidos grasos poliinsaturados de C20, como ácido araquidónico. Éstos participan en una diversidad de procesos fisiológicos, y también pueden mediar muchas respuestas potencialmente patológicas.
Las prostaglandinas son eicosanoides que tienen un anillo de ciclopentano. La prostaglandina E2 puede causar constricción de vasos sanguíneos, y el tromboxano A2 interviene en la formación de coágulos sanguíneos, que en algunos casos pueden bloquear el flujo de sangre al corazón o al cerebro. El leucotrieno D4, mediador de la contracción de músculos lisos, también provoca la constricción bronquial de los asmáticos. La aspirina (ácido acetilsalicílico) alivia el dolor, la fiebre, la hinchazón y la inflamación, al inhibir la síntesis de las prostaglandinas.

Las membranas biológicas están formadas por bicapas lipídicas y proteínas
Las membranas biológicas definen los límites externos de las células, y separan compartimientos dentro de ellas. Son componentes esenciales de todas las células vivas. Una membrana típica está formada por dos capas de moléculas de lípidos y muchas proteínas embebidas en ella.
Algunas proteínas contenidas en las membranas sirven como bombas selectivas que controlan en forma estricta el transporte de iones y de moléculas pequeñas que entran y salen de la célula. Las membranas también son responsables de generar y mantener la concentración de gradientes de protones, esenciales para la producción de ATP. Los receptores en las membranas reconocen señales extracelulares y las comunican al interior de la célula.
Bicapas lipídicas
Los glicerofosfolipidos y los glicoesfingolipidos anfipáticas pueden formar monocapas bajo ciertas condiciones lo que los hace ideal para formar bicapa lipídicas.
Las bicapas lipídicas son el principal componente estructural de todas las membranas biológicas, incluyendo membranas plasmáticas y membranas internas de eucariotas.
Las interacciones no covalente entre las moléculas de lípidos en las bicapas hace que la membrana sea flexible y les permite autosellarse.
Las bicapas lipídicas son el principal componente estructural de todas las membranas biológicas, incluyendo membranas plasmáticas y membranas internas de eucariotas.
Las interacciones no covalente entre las moléculas de lípidos en las bicapas hace que la membrana sea flexible y les permite autosellarse.

Modelo fluido de mosaico para membranas biológicas
Una membrana biológica típica contiene de un 25 a un 50% de lípidos, y de un 50 a un 75% de proteínas, en masa, con menos de 10% de carbohidratos como componente de glicolípidos y glicoproteínas. Los lípidos son una mezcla compleja de fosfolípidos, glicoesfingolípidos (en animales) y colesterol (en algunos eucariotas). El colesterol y algunos otros lípidos que por sí no forman bicapas (30% del total) están estabilizados en el arreglo de bicapa por el otro 70% de los lípidos en la membrana. Las composiciones
de las membranas biológicas varían en forma considerable entre las especies, y aun entre distintos tipos celulares en organismos multicelulares.
de las membranas biológicas varían en forma considerable entre las especies, y aun entre distintos tipos celulares en organismos multicelulares.
Una membrana biológica es más gruesa que una bicapa lipídica: en forma típica tiene de 6 a 10 nm de espesor. S. Jonathan Singer y Garth L. Nicolson propusieron en 1972 el modelo de mosaico fluido y todavía tiene validez general para describir el arreglo de lípidos y proteínas dentro de una membrana. Según el modelo del mosaico fluido, la membrana es una estructura dinámica en la que se pueden difundir lateralmente o girar dentro de la bicapa, en forma rápida y aleatoria, las proteínas y los lípidos. Las proteínas de membrana se conciben como témpanos de hielo flotando en un mar muy fluido de bicapa lipídica.

Las bicapas lipídicas y las membranas son estructuras dinámicas
Los lípidos en una bicapa están en movimiento constante, dando a las bicapas lipídicas muchas de las propiedades de los fluidos. Los lípidos tienen varios tipos de movimiento molecular dentro de las bicapas:
a) Difusión lateral, es relativamente rápida y el movimiento de los lípidos dentro del plano de una monocapa. Es bidimensional.
b) Difusión transversal o basculante de los lípidos es muy lenta y es el paso de lípidos de una monocapa de la bicapa a la otra.
b) Difusión transversal o basculante de los lípidos es muy lenta y es el paso de lípidos de una monocapa de la bicapa a la otra.

Todas las células sintetizan membranas nuevas agregando lípidos y proteínas a sus membranas existente.
Al extenderse la membrana plasmática, la célula aumenta de tamaño.
Al extenderse la membrana plasmática, la célula aumenta de tamaño.
Al final la célula se dividirá y cada célula hija hereda una parte (por la mitad) de la membrana progenitora.
Difusión de proteínas en membranas
1970, L. D. Frye y Michael A., realizaron experimentos para probar si las proteínas de membrana se difunden dentro de la bicapa lipídica. Con microscopia de inmunofluorescencia, observaron que las proteínas marcadas se entrecruzaron dentro de 40 minutos después de la fusión celular. Al menos algunas proteínas de membrana se difunden libremente en las membranas biológicas.

Tres clases de proteínas de membrana
Las membranas celulares e intracelulares contienen proteínas
especializadas enlazadas en la membrana.
1. Proteínas integrales de membrana.
2. Proteínas especificas de membrana.
3. Proteínas de membranas ancladas a lípidos.
Según su modelo de asociación con la bicapa
lipídica:
Las membranas celulares e intracelulares contienen proteínas
especializadas enlazadas en la membrana.
1. Proteínas integrales de membrana.
2. Proteínas especificas de membrana.
3. Proteínas de membranas ancladas a lípidos.
Según su modelo de asociación con la bicapa
lipídica:
1. Proteínas integrales de membrana:
Estas proteínas se caracterizan por tener regione hidrofóbicas incrustadas en el interior hidrofóbico de la bicapa lipídica. Abarcan totalmente la bicapa, con una parte de la proteína expuesta sobre la superficie externa o otra expuesta sobre la superficie interna.

2. Proteínas especificas de membrana.
Se asocian a una cara de la membrana mediante interacciones de carga – carga y con puentes de hidrogeno, con las proteínas integrales de membrana o con los grupos de cabeza polar de los lípidos de membrana.

3. Proteínas de membranas ancladas a lípidos.
Están unidas a una membrana mediante un enlace covalente con un ancla lipídico.
Los tres tipos de anclas se pueden encontrar en la misma membrana, pero no forman un complejo.
a) Proteína anclada a un acilo graso.
b) Proteína de membrana anclada a un prenilo. Las proteínas de membranas ancladas en acilo graso y prenilo también existen en la hojilla citoplasmática (externa) de la membranas intracelulares.
c) Proteína anclada por glicosilfosfatidilinositol.
a) Proteína anclada a un acilo graso.
b) Proteína de membrana anclada a un prenilo. Las proteínas de membranas ancladas en acilo graso y prenilo también existen en la hojilla citoplasmática (externa) de la membranas intracelulares.
c) Proteína anclada por glicosilfosfatidilinositol.

Transporte de membrana
A.Termodinámica del transporte en la membrana
B.Poros y canales
C.Transporte pasivo
D.Transporte activo
E.Endocitosis y exocitosis
A.Termodinámica del transporte en la membrana
B.Poros y canales
C.Transporte pasivo
D.Transporte activo
E.Endocitosis y exocitosis
A.Termodinámica del transporte en la membrana
Para una molécula A, la concentración en el interior de la membrana es [Aint] y la concentración en el exterior es [Aext]. El cambio de energía libre de Gibbs asociado al transporte de moléculas de A es:

Si la concentración de A en el interior de la célula es mucho menor que fuera de la célula, entonces ΔG transporte será negativa y se favorecerá termodinámicamente la entrada de A a la célula. Por ejemplo, si [Aint] = 1 mM y [Aext] = 100 mM, entonces, a 25 °:

B. Poros y canales
Los poros y los canales son proteínas transmembranales (porinas) con un paso central para iones y moléculas pequeñas. (En general, el término poro se usa en las bacterias, y canal para los animales). En este proceso no se requiere energía. Se controla por difusión.

Transporte de membrana a través de un poro o un canal. Un pasaje central permite que las moléculas y iones del tamaño, carga y geometría adecuados atraviesen la membrana en cualquier dirección.
C.Transporte pasivo:
El transporte pasivo también se llama difusión facilitada, porque no requiere fuente de energía. La proteína de transporte acelera el movimiento del soluto a favor de su
gradiente de concentración. Las proteínas de transporte son parecidas a las enzimas, porque aumentan la velocidad de un proceso que es termodinámicamente favorable. La ecuación que describe esta dependencia se parece a la ecuación de Michaelis-Menten para catálisis enzimática.
gradiente de concentración. Las proteínas de transporte son parecidas a las enzimas, porque aumentan la velocidad de un proceso que es termodinámicamente favorable. La ecuación que describe esta dependencia se parece a la ecuación de Michaelis-Menten para catálisis enzimática.

Cinética del transporte pasivo. La velocidad inicial de transporte aumenta con la concentración del sustrato, hasta que se alcanza un máximo. Ktr es la concentración del sustrato a la cual la velocidad de transporte es la mitad de la máxima.
Tipos de transporte activo y pasivo
Los transportadores de membrana más simples, sean activos o pasivos, realizan uniporte; esto es, sólo llevan un solo tipo de soluto a través de la membrana . Muchos transportadores hacen el transporte simultáneo de dos moléculas de diferentes solutos. Si ambas moléculas se transportan en la misma dirección, el proceso se llama simporte. Si se transportan en direcciones opuestas, el proceso es antiporte

D.Transporte activo:
El transporte activo se parece al transporte pasivo en el mecanismo y propiedades cinéticas generales. En el transporte activo, el soluto se mueve contra un gradiente de concentración, contra una diferencia de carga, o ambas cosas. El transporte activo debe acoplarse a una reacción productora de energía para contrarrestar el cambio desfavorable de energía libre de Gibbs, para transporte sin ayuda.
El transporte activo primario está activado por una fuente directa de energía, como ATP o luz. Por ejemplo, la bacteriorrodopsina usa energía luminosa para generar un gradiente de concentración de protones a través de la membrana, que se puede usar para formación de ATP. El transporte activo secundario está impulsado por un gradiente de concentración. El transporte cuesta arriba activo de un soluto se acopla con el transporte cuesta abajo de un segundo soluto que estaba concentrado por el transporte activo primario.
El transporte activo primario está activado por una fuente directa de energía, como ATP o luz. Por ejemplo, la bacteriorrodopsina usa energía luminosa para generar un gradiente de concentración de protones a través de la membrana, que se puede usar para formación de ATP. El transporte activo secundario está impulsado por un gradiente de concentración. El transporte cuesta arriba activo de un soluto se acopla con el transporte cuesta abajo de un segundo soluto que estaba concentrado por el transporte activo primario.

E.Endocitosis y exocitosis
En las células eucariotas, aunque no en todas, las proteínas (y ciertas sustancias grandes) se mueven hacia adentro y afuera de la célula por endocitosis y exocitosis, respectivamente.
La endocitosis es el proceso mediante el cual las macromoléculas son rodeadas por la membrana plasmática, y son llevadas al interior de la célula dentro de una vesícula lipídica.
La exocitosis se parece a la endocitosis, pero la dirección del transporte es la inversa. Durante la exocitosis, los materiales destinados a ser secretados de la célula se encierran en vesículas mediante el aparato de Golgi. A continuación las vesículas se funden con la membrana plasmática y liberan su contenido al espacio extracelular.
La endocitosis es el proceso mediante el cual las macromoléculas son rodeadas por la membrana plasmática, y son llevadas al interior de la célula dentro de una vesícula lipídica.
La exocitosis se parece a la endocitosis, pero la dirección del transporte es la inversa. Durante la exocitosis, los materiales destinados a ser secretados de la célula se encierran en vesículas mediante el aparato de Golgi. A continuación las vesículas se funden con la membrana plasmática y liberan su contenido al espacio extracelular.

En este vídeo explica un poco sobre el transporte a través de la membrana
Fármacos relacionados con lípidos
Colestiramina (Questran)
Es una resina polimérica con capacidad
para fijar los ácidos biliares. Inicialmente, se utilizó para tratar el prurito
secundario a la colestasis, pero hoy día su principal indicación es el
tratamiento de la hipercolesterolemia con hipertrigliceridemia concomitante.
También se ha utilizado esta resina para tratar la enterocolitis producida por
el Clostridium difficile

Dosis:
Administración oral:
- Adultos:
inicialmente 4 g de colestiramina tres veces al día, antes de las comidas.
Las dosis de deben ajustar de acuerdo con la respuesta clínica. La dosis
de mantenimiento más usual es de 4 a 8 g de la resina, 2 a 4 veces al día,
con las comidas y a la hora de acostarse. La dosis máxima recomendada es
de 24 g/día como hipocolesterolemiante y de 16 g/día como antiprurítico
- Adolescentes:
la dosis de 8 g/día ha mostrado ser eficaz y segura en esta población. La
dosis de debe repartir en dos tomas, antes de las comidas
- Niños
de 6 a 12 años: se recomienda 80 mg/kg por vía oral o 2.35 g/m2 tres veces
al día con las comidas. Alternativamente pueden administrarse inicialmente
de 2 a 4 g dos veces al día antes de las comidas. Estas dosis se pueden
incrementar hasta conseguir la respuesta clínica adecuada o hasta el
desarrollo de efectos adversos intolerables, hasta un máximo de 8 g/día
- Niños de < 6 años: la seguridad y eficacia de la colestiramina no han sido evaluados en esta población

Mecanismo de acción
Actúa uniéndose a los ácidos biliares e impidiendo su
reabsorción. De esta manera se fomenta la transformación del colesterol
hepático en ácidos biliares. Secundariamente, la disminución del colesterol
incrementa la actividad de los receptores LDL de los hepatocitos, con lo que se
incrementa la eliminación del colesterol LDL plasmático.
Lovastatina (Mevacor)
Es un fármaco miembro de la familia de las estatinas, usado para disminuir el colesterol y prevenir enfermedades
cardiovasculares.

Vías de administración (formas de uso)
Oral.
 Dosis
Dosis
En adultos:
- 20 mg/día vía oral inicialmente, como dosis única con la
cena, aumentándolo en intervalos de 4 semanas o más hasta una dosis de mantenimiento
de 20-80 mg/día en dosis únicas o divididas. Los pacientes con niveles de
colesterol por encima de 300 mg/dL (7,8 mmol/L) con dieta como terapia pueden
ser iniciados con 40 mg/día.
Absorción
La absorción, al igual que la
mayoría de las estatinas, es limitada, reduciéndose a un máximo del 30%. Sufre
efecto de primer paso metabólico. Esto junto con la escasa absorción le da una
biodisponibilidad muy baja, en torno al 5%. Diversos estudios muestran que su
absorción en ayunas es de 2/3 a la registrada inmediatamente después de las
comidas.
Mecanismo
de acción
El β-hidroxiácido de
lovastatina es un inhibidor de la 3-hidroxi-3-metilglutaril-coenzima
A reductasa (HMG-CoA reductasa).
Esta enzima cataliza un paso esencial de la vía
del mevalonato, la conversión de la HMG-CoA amevalonato, que es un metabolito
clave en la en la biosíntesis de colesterol.
En el esquema adjunto puede observarse el nivel de bloqueo de las estatinas así
como de otras sustancias en la biosíntesis del colesterol. La inhibición de la
enzima se realiza de forma competitiva. El bloqueo de la síntesis hepática del
colesterol produce una activación de las proteínas reguladoras SREBP (sterol regulatory elements-binding
proteins), que activan la transcripción de proteínas y, por tanto, producen
una mayor expresión del gen del receptor de LDL y un aumento en la cantidad de
receptores funcionales en el hepatocito.

Dosis
20-40 mg/día, por la noche, ajuste de dosis
cada 4 sem; máx. 80 mg/día. Niños > 9 años: 20-40 mg/día, por la noche,
ajuste de dosis cada 6 sem; máx. 80 mg. Enf. cardiaca coronaria después de
intervención coronaria: 80 mg/día.
Mecanismo de
acción
 La fluvastatina interfiere con la
actividad de la hidroximetilglutaril-coenzima A reductasa (HMG-CoA), una enzima
hepática, con lo que se interrumpe la vía sintética de la biosíntesis de
colesterol en el hombre. Como consecuencia, los niveles del colesterol hepático
son reducidos, estimulándose la captación de las LDLs. La fluvastatina reduce
el colesterol total circulante, el colesterol asociado a las LDLs y los
triglicéridos. La fluvastatina es más potente que la lovastatina como inhibidor
de la HMG-CoA reductasa.
La fluvastatina interfiere con la
actividad de la hidroximetilglutaril-coenzima A reductasa (HMG-CoA), una enzima
hepática, con lo que se interrumpe la vía sintética de la biosíntesis de
colesterol en el hombre. Como consecuencia, los niveles del colesterol hepático
son reducidos, estimulándose la captación de las LDLs. La fluvastatina reduce
el colesterol total circulante, el colesterol asociado a las LDLs y los
triglicéridos. La fluvastatina es más potente que la lovastatina como inhibidor
de la HMG-CoA reductasa.
What's On Casino? - Casinoworld
ResponderBorrarCasinoworld welcomes you 카지노사이트 to the home of a casino, 바카라 사이트 where our players can play their favourite games at the leading online casino.Casino Games 온카지노 · Casinos · Why Play · How to Start Playing at